1. INTRODUCCIÓN
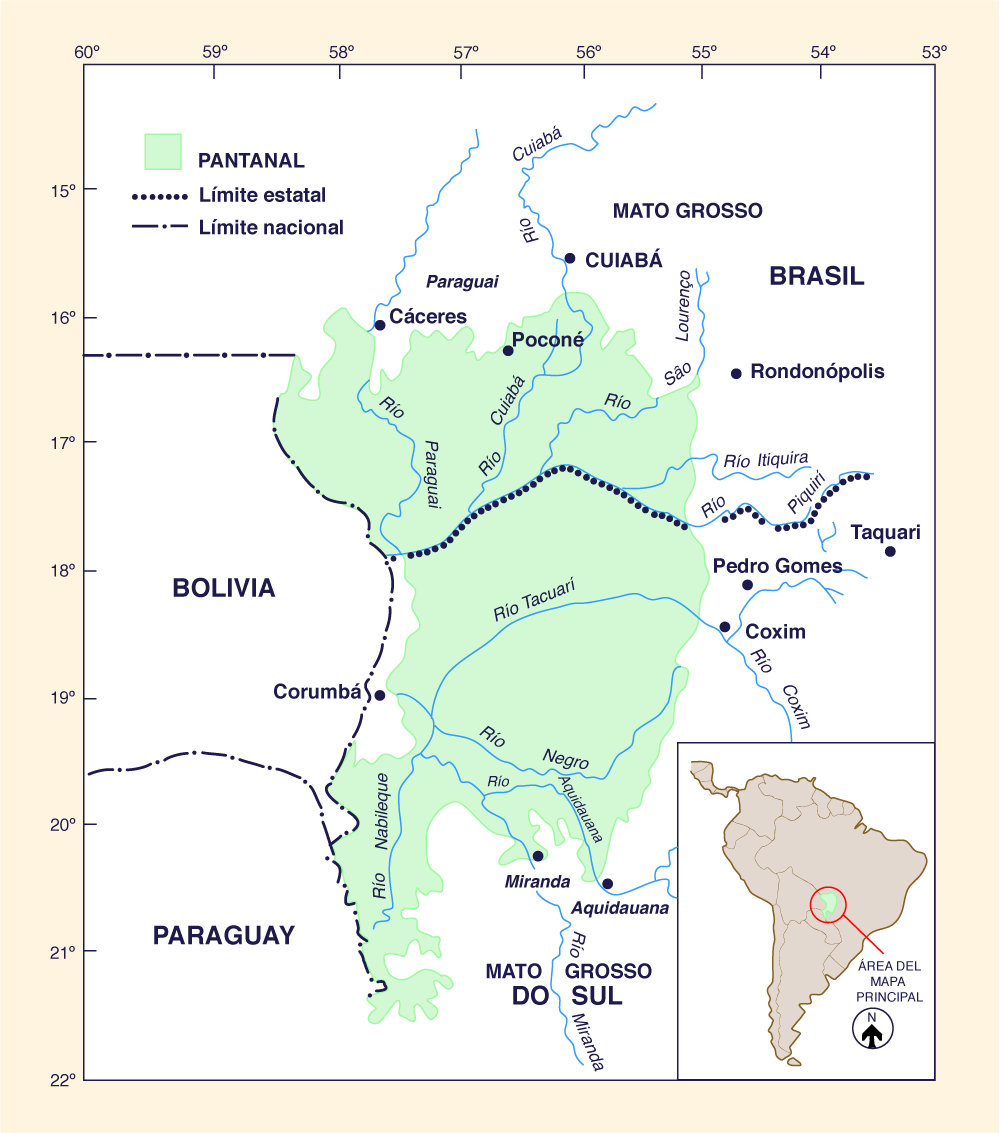
El Pantanal do Mato Grosso es uno de los ecosistemas de humedales tropicales más grandes del mundo, abarcando un área de aproximadamente 140,000 km2 en la cuenca alta del Río Paraguay, en el centro-oeste del Brasil (Fig. 1). "Pantanal" en portugués significa gran pantano, lo que implica que la región está sujeta a inundaciones periódicas extensas. Sin embargo, grandes regiones del Pantanal se inundan sólo durante la cresta anual del Río Alto Paraguay y sus afluentes, y gran parte de tierra firme no inundada se distribuye por toda la región.
El área es plana a ligeramente ondulada, y la mayor parte se encuentra a unos 100 m sobre el nivel medio del mar. La mezcla de pantano permanente, pantano estacional y tierra firme, así como la contigüidad del Pantanal a otros importantes ecosistemas de la América del Sur (la selva amazónica al norte y noroeste, las sabanas tropicales del centro de Brasil al este, y la región seca o bosque de tipo chaco de la vecina Bolivia al suroeste), ha contribuido a la riqueza y variedad de su vegetación. En la literatura existente, la vegetación a menudo se marca como una sola unidad y se la denomina "complejo del Pantanal". En realidad, este complejo es un mosaico de varias comunidades diferentes, con frecuentes cambios abruptos generalmente correlacionados con la topografía, y con muchos ecotonos (Prance y Schaller, 1982).
Una característica geomorfológica peculiar del paisaje del Pantanal son los montículos con vegetación (de varios tamaños y formas, y de planta aproximadamente circular o elíptica) que existen en la gran extensión de pastizales inundados estacionalmente. En el Brasil se han utilizado varios términos para describir estos curiosos montículos de tierra: murundú, covoal, morrote, cocuruto, moncheio, capãozinho, e ilha (Oliveira Filho y Furley, 1990). Los montículos de tierra más grandes, que pueden alcanzar diámetros de más de 100 m, se han denominado capões y se han distinguido de los capões de aterro o aterros de bugre, que son decididamente artificiales (Cunha, 1990).
En las últimas tres décadas han aparecido en la literatura de lengua inglesa relatos breves de estos montículos brasileños (Cole, 1960; Askew et. al., 1970; Ratter et. al., 1973; Eiten, 1975; Prance y Schaller, 1982; Furley y Ratter, 1988). Estos relatos atribuyen la génesis y evolución de los montículos a factores abióticos y bióticos; sin embargo, queda por dilucidar el preciso papel de estos mecanismos. A diferencia de los pastizales abiertos adyacentes que no apoyan especies vegetativas leñosas, los montículos de tierra del Pantanal presentan vegetación leñosa, lo que revela su estrecha asociación con los bosques de sabana o cerrados del centro del Brasil. En este artículo se han revisado los conocimientos actuales sobre los montículos con vegetación de las sabanas tropicales del centro de Brasil. La naturaleza y la capacidad de supervivencia del hábitat del montículo se explica en términos de la competencia entre las comunidades de plantas de cerrado y de campo. Se discuten varias clasificaciones de montículos y se desarrolla una clasificación tentativa basada en la escala. 2. REVISIÓN DE LITERATURA Los montículos de tierra brasileños se han mencionado brevemente en la literatura que trata de la vegetación del centro de Brasil. Cole (1960) señaló: "A través de los pastizales hay grupos de árboles, generalmente la especie de cerrado ipê roxa, ipê amarelo (ambas Tecomia spp.) y el lixeiro, este último generalmente creciendo cerca de los nidos de termitas alrededor de los cuales se acumulan los sedimentos, permitiendo proteger pequeñas porciones del suelo contra las crecidas. Lejos de los ríos, en las áreas por encima del nivel de las crecidas normales, los grupos de árboles se vuelven más numerosos, y la transición se produce del pantanal bajo al alto. Estos últimos transicionan a campo cerrado, donde la ausencia total de inundaciones se asocia con una distribución aparentemente aleatoria de árboles". En un estudio de suelos del noreste del Mato Grosso, Askew et. al. (1970) notaron que los campos se encuentran en los valles (donde el suelo es más húmedo y generalmente saturado hasta la superficie durante la temporada de lluvias), y que muchas áreas de campo contienen islas elevadas de cerrado que poseen termitaria. En un estudio relacionado, Ratter et. al. (1973) describieron un tipo de "sabana de termitas" en la que montículos de hasta 2-3 m de altura y no más de 6-7 m de diámetro, cada uno con un grupo de árboles de cerrado y una o más termitaria, están esparcidos de manera notablemente uniforme sobre un pastizal plano, sin árboles, que se inunda en la estación húmeda pero está seca desde mayo o junio hasta diciembre o enero. Ratter et. al. (1973) estudiaron tales sabanas de termitas a pocos kilómetros al sureste de Xavantina, y reportaron haber visto áreas mucho más extensas en el valle del rio das Mortes y en el valle del río Araguaya, cerca de la Ilha do Bananal.
Eiten (1975), refiriéndose a los montículos de tierra de la Serra do Roncador (noreste del Mato Grosso), señaló que en casi todos los campos del lado de los valles, excepto aquéllos en laderas empinadas, había arboledas circulares dispersas del matorral cerrado de varios metros de diámetro. Estas arboledas siempre se encuentran sobre un suelo ligeramente elevado por encima del nivel general del campo, desde aproximadamente 0.3 m a un poco más de 1 m. Encima de cada una de estas plataformas de suelo elevado hay un sólo nido de termitas de 1.0 a 1.5 m de altura. El suelo de las plataformas es siempre del mismo gris claro que el subsuelo del campo, al igual que el nido mismo. Aparentemente, las termitas han construido esta plataforma para evitar que la base de su nido toque el suelo saturado. Eiten afirmó que nunca vio una plataforma sin un nido bien formado, ni vio un nido degradado en una plataforma, situación que le llevó a observar que las plataformas no son el resultado de la degradación de los nidos. Además, observó que la cantidad de suelo en la plataforma es muchas veces mayor que la de un nido y no es tan duro como el suelo del nido. Prance y Schaller (1982) notaron el fuerte elemento de cerrado en el Pantanal do Mato Grosso, particularmente en la parte oriental. Estos cerrados están dominados por especies como Bowdichia virgiloides, Caryocar brasiliense, Curatella americana y Qualea parviflora, todas ellas típicas de los cerrados del planalto del Brasil Central. Observaron que el cerrado ocurre principalmente en las tierras altas no inundadas, pero también hacia el borde oriental del Pantanal, donde la tierra se inunda sólo por períodos cortos en el apogeo de la temporada de inundaciones. Tal cerrado húmedo tiende a consistir en numerosas islas de cerradão (bosque denso) en áreas ligeramente elevadas que no están inundadas. Numerosas islas pequeñas de cerrado rodean las colinas de termitas y la actividad de las termitas aumenta el tamaño de estas islas, incrementando la elevación. Araujo Neto et. al. (1986) informaron sobre los campos de pequeños montículos o campos de murundús de Fazenda Agua Limpa, en el Distrito Federal (Brasilia). Estos murundús son casi elípticos en planta, con dimensiones de longitud promedio de 7.0 × 5.5 m, altura de 0.05 m a más de 2 m, volumen de 0.1 m3 a 141.5 m3 y densidad aéreal de 26 a 61 ha-1. Los campos de montículos consisten en patrones repetidos de montículos redondeados, cubiertos por plantas leñosas de la sabana, y depresiones vecinas dominadas por pastos, juncos y otra vegetación pequeña. Estos campos de montículos de tierra logran su desarrollo más espectacular entre las formaciones superiores y más secas (campo cerrado) y las formaciones del valle húmedo inferior del bosque de galería (mata ciliar). Se cree ampliamente que los murundús son montículos de termitas, análogos a las sabanas de termitas del centro-sur de África. En el centro del Brasil, esta creencia no se basa en mediciones de campo detalladas, sino en la observación general de que muchos montículos están ocupados por termitas. Por lo tanto, no está claro si las termitas construyen activamente todo o parte de los montículos, o si simplemente ocupan los montículos, donde las condiciones ambientales son más adecuadas. Araujo Neto et. al. (1986) concluyeron que muchos de los montículos no estaban ocupados por termitas y que no se ha podido discernir ninguna evidencia pedológica que indique una ocupación pasada. Interpretaron los montículos como productos residuales de la erosión diferencial y supusieron que la escorrentía superficial, los efectos de la filtración y el agua subterránea fueron los procesos dominantes en la génesis de los murundús. Sin embargo, notaron que la mayoría de los montículos más grandes mostraban signos de actividad de termitas. Furley y Ratter (1988) observaron que las especies de árboles de cerrado que son más resistentes al anegamiento (por ejemplo, Curatella americana y Byrsonima crassifolia), son comunes cerca del límite cerrado/campo y en islas elevadas de tierra en campos húmedos. La distribución de estas islas produce los campos de murundús, que consisten en una extensión de campos húmedos salpicados con un patrón regular de montículos de tierra elevados, con árboles de cerrado, arbustos y, a menudo, termitaria. Cunha (1990) describió los capões de cerrado en el Pantanal de Poconé, una de las varias subregiones que constituyen el Pantanal do Mato Grosso. Estos montículos de tierra son de forma circular a elíptica, de 4-7 m de diámetro, y tienden a estar dominados por el lixeiro (Curatella americana). Cunha (1990) también ha documentado capões mucho más grandes, típicamente de forma elíptica, de longitudes de hasta 300 m, anchos de hasta 80 m, y escasamente distribuidos alrededor de los campos sujetos a inundaciones estacionales. Los aterros de bugre son diques construidos por los moradores indígenas de la región, aparentemente para proporcionar un área de terreno elevado para protección durante la temporada anual de inundación. Las excavaciones superficiales realizadas en estos aterros de bugre invariablemente revelan restos arqueológicos (fragmentos de cerámica y demás bienes parafernales), lo que confirma el origen geológicamente reciente y artificial de estos rasgos geomorfológicos.
3. LA NATURALEZA DE LOS MONTÍCULOS DE TIERRA
El factor abiótico: Geomorfología El papel del factor abiótico se hipotetiza de la siguiente manera: después del inicio, debido a la superficie u otras irregularidades, los montículos de tierra incipientes son fácilmente colonizados por árboles y arbustos de cerrado, particularmente por aquellas especies que son más tolerantes al anegamiento. El establecimiento de vegetación en los montículos actúa para reducir la velocidad de las inundaciones estacionales a su alrededor, contribuyendo aún más (por sedimentación diferencial) a un aumento de su tamaño. Durante los años extremadamente húmedos, se puede conjeturar que los montículos serán cubiertos por las inundaciones, aunque solo por períodos breves. Como la reducción de la velocidad de las aguas de la inundación es gradual desde el perímetro hacia el centro de cada montículo, siendo más marcado en el centro, se prevé el proceso de acumulación gradual del montículo por sedimentación diferencial a lo largo de milenios. El factor abiótico no excluye la actividad de las termitas, que pueden optar por aprovechar el ambiente más seco del montículo de tierra. El tamaño y la forma de los montículos más grandes (capões), junto con las observaciones de campo que documentan la presencia limitada o incluso la ausencia de termitas, apoyan la hipótesis de la sedimentación diferencial. El factor biótico: Termitaria
Mathews (1977) ha desarrollado una teoría para explicar el papel de termitaria en la génesis de los montículos de tierra brasileños. Sugirió que los montículos más pequeños (murundús) se forman a través de un largo proceso que involucra diferentes etapas, cada etapa corresponde a la acción de una especie de termitas diferente, como sigue: (1) Inicialmente, una especie más tolerante a los suelos húmedos (Armitermes cerradoensis) coloniza el campo durante la estación seca; (2) durante la temporada de lluvias, otra especie de termita (Anoplotermes sp.) busca refugio alrededor del nido de A. cerradoensis, aportando larvas y suelo al montículo; (3) tras la muerte de la colonia de En un estudio reciente de un campo de monchões en una depresión inundada estacionalmente en las cercanías de Cuiabá (Mato Grosso), Oliveira Filho (1988) encontró evidencia que indica que los suelos de los montículos de tierra probablemente se formaron a través de la degradación gradual y erosión de sucesivas generaciones de termitaria. Esto lo llevó a proponer una teoría para explicar la formación de los monchões en términos de un proceso de sucesión primaria en el que las islas ecológicas (es decir, los monchões) crecen gradualmente en tamaño y, en consecuencia, en complejidad, diversidad de especies, biomasa, y energía total.
4. LA COMPETENCIA ENTRE CERRADO Y CAMPO
Hay tres tipos de vegetación fisonómica y florísticamente distintos que pueblan las llanuras altamente disecadas del noreste del Mato Grosso: (1) cerrado (incluida su variante cerradão), Eiten (1975) ha documentado la nitidez del límite cerrado/campo. Dentro de 1 m, o incluso 0.5 m, el cambio de los arbustos y árboles bajos del cerrado a la capa de hierba sin plantas leñosas del campo es completo. La razón por la que el cerrado se detiene repentinamente y da paso al campo parece ser que las plantas del cerrado no pueden establecerse a partir de semillas en un suelo continuamente húmedo. En general, el campo ocupa un sitio con un nivel freático más bajo y más fluctuante, mientras que el cerrado ocupa un terreno más alto donde el suelo rara vez o nunca permanece saturado. Por lo tanto, el límite cerrado/campo claramente definido se atribuye al anegamiento en sí, y no al cambio de condición química (y color) del suelo, que es el resultado del anegamiento. En casi todos los casos, el cerrado se detiene repentinamente al borde del campo, aparentemente debido a la competencia entre los dos tipos de vegetación como comunidades de plantas. Tan pronto como la superficie del suelo es elevada por las termitas y permanece permanentemente no saturada y la superficie puede secarse durante la estación seca, las plantas del cerrado lo invaden con toda su fuerza. En la mayoría de los lugares, el nivel del agua subterránea tiene una relación muy precisa con el límite de cerrado/campo. La relación entre el nivel del agua subterránea y el límite se puede explicar en términos de los altos niveles de agua en la temporada de lluvias. Las especies de cerrado tolerantes al anegamiento pueden crecer en campos abiertos en lugares cercanos al límite del cerrado donde el nivel del suelo es solo unos centímetros más alto que en otros lugares. La observación de que las islas más grandes en el paisaje pantanal están densamente cubiertas con vegetación de cerrado apoya la opinión de que el nivel del agua subterránea ejerce un control preciso sobre el límite de/ cerrado/campo. La fluctuación estacional en la profundidad del nivel freático es de importancia crucial para la vegetación de dos maneras: (1) al influir en la capacidad de las plantas para absorber agua, especialmente en años de sequía; y (2) restringiendo la actividad de las raíces en la estación húmeda a las capas comparativamente bien oxigenadas del suelo, a excepción de la vegetación del bosque en galería que tiene sistemas de raíces especialmente adaptados a las condiciones anaeróbicas y anegadas. En tales condiciones, diferencias muy pequeñas en la topografía o el suelo son suficientes para inclinar la balanza a favor de un tipo de vegetación u otro. La humedad del suelo está altamente correlacionada con la principal tendencia de variación vegetacional. La falta patente de árboles en los campos húmedos es sorprendente, particularmente porque una amplia gama de especies leñosas coloniza con éxito tanto los interfluvios (que son más secos que los campos) como los lados de los arroyos (que son más húmedos). La ausencia de especies leñosas altas en áreas que son intermedias en sus características físicas puede deberse a la naturaleza fluctuante del nivel freático y la consiguiente humedad del suelo. Por lo tanto, los árboles parecen ser capaces de tolerar tanto condiciones permanentemente húmedas (bosque de galería) como secas (cerrado), pero no una alternancia extrema de saturación y desecación (Goldsmith, 1974).
5. CLASIFICACIÓN DE LOS MONTES DE TIERRA
Los montículos de tierra brasileños varían en tamaño, forma, orientación, cobertura vegetal, propiedades del suelo, actividad de las termitas, escorrentía superficial y agua subterránea. Oliveira Filho y Furley (1990) han intentado recientemente una clasificación de los murundús basada en procesos formativos. Observaron que el rol de las termitas parecía tener aproximadamente la misma importancia que el de los procesos hidrológicos/geomorfológicos, particularmente con respecto a los montículos más pequeños (murundús). Esto los llevó a clasificar los montículos en: (1) aquéllos formados principalmente por procesos abióticos, es decir, procesos hidrológicos/geomorfológicos, incluyendo el efecto de inundaciones, escorrentías superficiales y aguas subterráneas; (2) aquéllos formados principalmente por procesos bióticos, es decir, por la actividad de las termitas, y (3) aquéllos de génesis mixta abiótica/biótica, con acción combinada de procesos hidrológicos/geomorfológicos y termitaria. Oliveira Filho y Furley (1990) observaron que la escorrentía superficial, la cual conduce a la erosión diferencial, parece ser un proceso dominante en la formación y desarrollo de algunos murundús, particularmente aquéllos que no han sido afectados por el agua subterránea. Supusieron que un proceso de erosión y sedimentación diferencial bien podría explicar la existencia de los montículos de tierra más grandes o capões encontrados en el bajo Pantanal del Mato Grosso. Sin embargo, parece más probable que los montículos más pequeños, o murundús de pantanal, hayan sido causados por la acción combinada de procesos hidrológicos/geomorfológicos y termitaria. La marcada diferencia de escala de murundús y capões sugiere una clasificación tentativa de los montículos sobre esta base. De acuerdo con este concepto, los montículos de tierra se clasifican en: (1) micromontículos, o murundús, y (2) macromontículos, o capões. Los micromontículos a menudo pueden mostrar una notable regularidad espacial cuando se observan desde el aire, particularmente aquéllos que tienen una influencia biótica importante. Smith (1971) ha mostrado una vista aérea de las llanuras aluviales del río Araguaya, donde es difícil explicar la precisa regularidad espacial de los pequeños montículos de tierra. Mathews (1977) ha explicado la regularidad espacial en términos de los dominios territoriales de las colonias de termitas que son hostiles entre sí.
Los micromontículos o murundús son típicamente los estudiados por Araujo Neto et. al. (1986) en Fazenda Agua Limpa, Brasilia, aunque también ocurren en otras regiones del centro de Brasil, incluido el noroeste del Mato Grosso, el oeste de Goiás y en el alto Pantanal del Mato Grosso.
6. SÍNTESIS El Pantanal del Mato Grosso y otros ecosistemas del centro de Brasil se encuentran dentro de la región climática del cerrado, un bioma tipo sabana que se caracteriza por su adaptación a una larga estación seca, la cual a menudo dura hasta siete meses. Son posibles varias instancias de rodales de cerrado, siendo la profundidad del nivel freático el factor ambiental crítico que determina la variación fisionómica y florística. La secuencia topográfica cerrado/campo/bosque de galería es común en muchas áreas del Mato Grosso. La vegetación del cerrado requiere un suelo bien drenado, sin acumulación de agua a largo plazo durante la temporada de lluvias. Por el contrario, los árboles de un bosque de galería tienen sistemas de raíces especialmente adaptados a un drenaje deficiente y al consecuente anegamiento.
Los campos de pastizales, situados entre el cerrado y el bosque de galería, son el único tipo de vegetación particularmente bien adaptados a alternancias extremas de saturación y desecación.
La naturaleza de los montículos de tierra se explica en términos de la competencia entre dos comunidades vegetales, en este caso, el cerrado, endémico de las sabanas del centro de Brasil, y el campo, cuyos pastos se adaptan mejor a alternancias extremas de saturación y desecación.
BIBLIOGRAFÍA Araujo Neto, M. D., P. A. Furley, M. Haridasan, y C. F. Johnson. 1986. The murundús of the cerrado region of Central Brazil. J. Trop. Ecol. 2, 17-35.
Askew, G.P., D. J. Moffatt, R. F. Montgomery, y P. L. Searl. 1970. Soil landscapes in northeastern Mato Grosso. Geogrl. J. 136,211-227.
Cole, M. M. 1960. Cerrado, caatinga and pantanal: the distribution and origin of the savanna vegetation of Brazil.
Geogrl J. 126, 16,179.
Cunha, C. N. 1990. Estudos florísticos e fitofisionómicos das principais formações arbóreas do Pantanal de Poconé-MT.
Master's thesis, Universidade Estadual de Campinas, São Paulo, Brazil.
Eiten. G. 1975. The vegetation of the Serra do Roncador. Biotrop. 7, 112-135.
Farley. P. A. 1986. Classification and distribution of murundús in the cerrado of Central Brazil. J. Biogeogr. 13, 265-268.
Farley, P. A. y J. A. Ratter. 1988. Soil resources and plant communities of the central Brazilian cerrado and their development. J. Biogeogr. 15, 97-108.
Goldsmith, F. B. 1974. Multivariate analysis of tropical grassland communities in Mato Grosso, Brazil. J. Biogeogr. 1, 111-122.
Mathews, A. G. A. 1977. Studies on termites from the Mato Grosso State. Brazil. Academia Brasileira de Ciências, Rio de Janeiro.
Oliveira Filho, A. T. 1988. A vegetação de um campo de monchões-microrrelevos associados a cupins--na regiã de Cuiabá (MT). Ph.D. dissertation, Universidade Fstadual de Campinas, São Paulo, Brazil.
Oliveira Filho, A. T. 1992. Floodplain 'murundús' of Central Brazil: evidence for the termite-origin hypothesis. J. trop. Ecol. 8, 1-19.
Oliveira Filho, A. T. y P. A. Farley. 1990. Monchão,
cocuruto, murundu. Ciência Hoje. 11, 30-37.
Prance, G. T. y Schaller, G. B. 1982. Preliminary study of some vegetation types of the Pantanal, Mato Grosso, Brazil. Brittonia, 34, 228-251.
Ratter, J. A., P. W. Richards, G. Argent, y D. R. Gifford. 1973. Observations of the vegetation of northeastern Mato Grosso I. The woody vegetation types of the Xavantina-Cachimbo expedition area. Phil. Trans. R. Soc. 266B, 449-492.
Smith, A. 1971. Mato Grosso: Last virgin land. Dutton. New York.
|


| 220102 |
| Documents in Portable Document Format (PDF) require Adobe Acrobat Reader 5.0 or higher to view; download Adobe Acrobat Reader. |